この章で学ぶこと
第 3 章で学んだ代謝の主役は 酵素 = タンパク質 でした。 そのタンパク質の 設計図 が DNA です。 第 4 章では、 遺伝がどのように親から子へ伝わり、 DNAがどのようにタンパク質を作るかを学びます。
- メンデルの法則 (優性・分離・独立) を説明できる
- DNAの二重らせん構造 (ワトソン・クリック) を理解する
- 半保存的複製 (メセルソン・スタール) のしくみを言える
- セントラルドグマ (DNA → RNA → タンパク質) を説明できる
- 転写・翻訳の流れとコドンを理解する
- 突然変異 の種類を言える
ポイント: 遺伝の研究史は 「現象 (メンデル) → 染色体 (サットン・モーガン) → 分子 (ワトソン・クリック) → 機能 (転写・翻訳)」 と進みました。 この流れを押さえると全体が見えます。
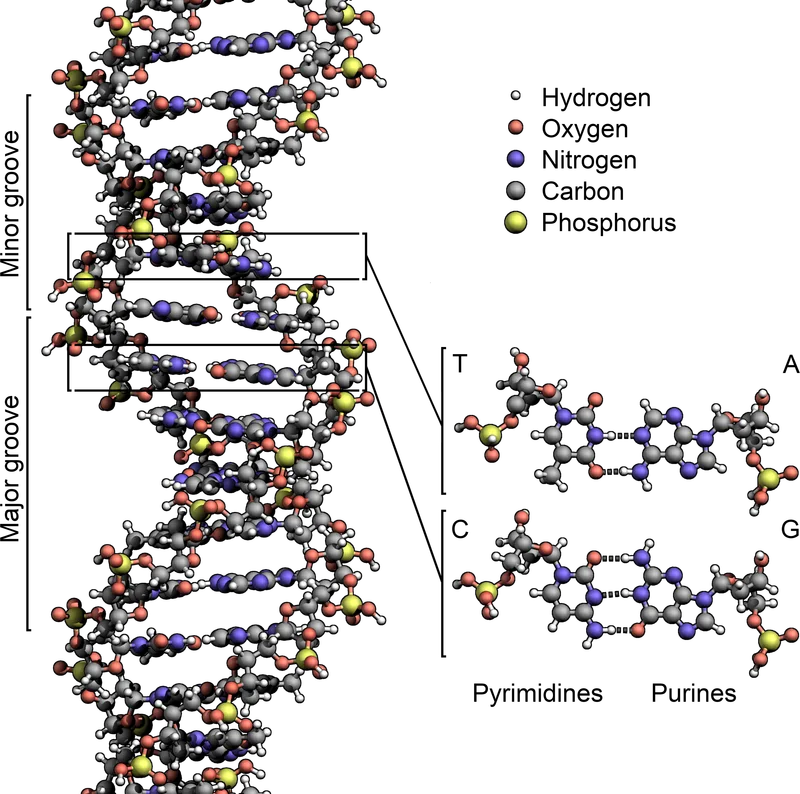 DNA の二重らせん構造 (1953 年ワトソン・クリックが提案)。 4 種類の 塩基 (A・T・G・C) が 相補的 に対 (A-T, G-C) を作り、 親鎖の配列がそのまま子に伝わる。
DNA の二重らせん構造 (1953 年ワトソン・クリックが提案)。 4 種類の 塩基 (A・T・G・C) が 相補的 に対 (A-T, G-C) を作り、 親鎖の配列がそのまま子に伝わる。
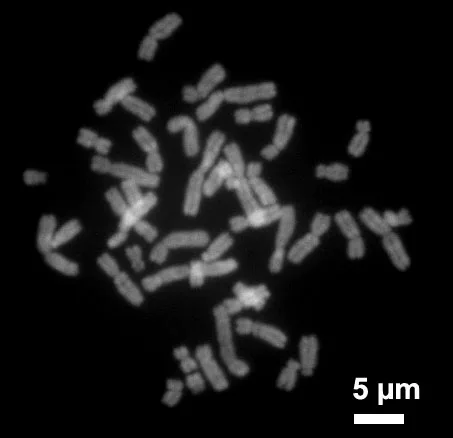 ヒト の 染色体 (蛍光染色)。 体細胞には 46 本 (23 対) あり、 そのうち 1 対が 性染色体 (XX = 女、 XY = 男)。 各染色体は 長いDNAがタンパク質と巻かれて凝縮した もの。
ヒト の 染色体 (蛍光染色)。 体細胞には 46 本 (23 対) あり、 そのうち 1 対が 性染色体 (XX = 女、 XY = 男)。 各染色体は 長いDNAがタンパク質と巻かれて凝縮した もの。
1. メンデルの遺伝法則
オーストリアの修道士 メンデル (Gregor Mendel、 1822-1884) が、 エンドウを 8 年間育てて統計的に解析し、 1865 年に発表した法則。 彼の研究は当時注目されず、 1900 年に 3 人の学者 (ド・フリース・コレンス・チェルマック) が同時に 「再発見」 したことで広まりました。
用語の整理
| 用語 | 意味 |
|---|
| 遺伝子 | 形質を決める因子 (現在ではDNAの 1 領域) |
| 対立遺伝子 (アレル) | 同じ場所に入る異なる型の遺伝子 (例: 丸を決める A としわを決める a) |
| ホモ接合 | 両方の対立遺伝子が同じ (AA・aa) |
| ヘテロ接合 | 両方の対立遺伝子が違う (Aa) |
| 顕性 (優性) | 雑種で表に出る形質 (大文字 A) |
| 潜性 (劣性) | 雑種で隠れる形質 (小文字 a) |
| 表現型 | 実際に見える形質 (例: 丸・しわ) |
| 遺伝子型 | 遺伝子の組み合わせ (AA・Aa・aa) |
大事: 「優性」 「劣性」 は強い・弱いという意味ではなく 「雑種で 表に出やすいかどうか」 だけを意味します。 紛らわしいため、 現在は 顕性・潜性 と呼ぶことが推奨されています。
メンデルの 3 法則
① 優性の法則 (顕性の法則)
ホモ接合の親 (AA × aa) を交配すると、 雑種第 1 代 (F₁) はすべて ヘテロ接合 (Aa) で 顕性形質だけ を示す。
② 分離の法則
F₁ (Aa) が配偶子 (花粉・卵細胞) を作るとき、 対立遺伝子が 1 つずつ分かれて 別の配偶子に入る。 結果、 F₁ 同士を自家受粉した F₂ では 表現型 = 顕性:潜性 = 3:1 になる。
③ 独立の法則
2 つの異なる対立遺伝子 (例: 丸 / しわ × 黄色 / 緑) は 互いに影響せず独立に配偶子へ分かれる。 結果、 F₂ で 9:3:3:1 の比が出る。 (※ ただしこの法則は 同じ染色体上にない遺伝子 に限る)
F2 の比 — 簡単な計算
| 親の組合せ | F₁ | F₂ (表現型) | 比 |
|---|
| AA × aa | Aa | A_:aa | 3:1 |
| AABB × aabb | AaBb | (A_B_):(A_bb):(aaB_):(aabb) | 9:3:3:1 |
2. 染色体と遺伝子
染色体説
メンデルの死後、 細胞学が進み、 1903 年にサットンが 「遺伝子は染色体に載っている」 と提案 (染色体説)。 1910 年にモーガンがショウジョウバエの実験で確認し、 「連鎖と組換え」 を発見 (ノーベル賞)。
連鎖と組換え
| 用語 | 意味 |
|---|
| 連鎖 | 同じ染色体上の遺伝子が 一緒に行動 すること |
| 組換え | 減数分裂時に相同染色体間で一部が入れ替わる (= 「乗換え」) |
| 組換え価 | 全配偶子中の組換え配偶子の割合 (%) |
組換え価が高いほど遺伝子の距離が遠いことを意味し、 これで 染色体地図 が作れます。
3. DNA — 遺伝物質の正体
DNA が遺伝物質だと分かるまで
| 年 | 研究 | 結果 |
|---|
| 1869 | ミーシャー | 細胞核から ヌクレイン を抽出 |
| 1928 | グリフィス | 肺炎双球菌の 形質転換 を発見 |
| 1944 | エイブリーら | 形質転換の正体が DNA と確定 |
| 1952 | ハーシー・チェイス | バクテリオファージ実験で DNA が遺伝物質と確定 |
| 1953 | ワトソン・クリック | DNA の 二重らせん構造 を発表 (1962 ノーベル賞) |
DNA の構造
DNA (Deoxyribo Nucleic Acid、 デオキシリボ核酸) は、 ヌクレオチド が多数つながった高分子。
1 つのヌクレオチド:
- リン酸 + デオキシリボース (糖) + 塩基 (4 種のうち 1 つ)
4 種の塩基:
| 種類 | 略 | 分類 | 相補ペア |
|---|
| アデニン | A | プリン | T |
| チミン | T | ピリミジン | A |
| グアニン | G | プリン | C |
| シトシン | C | ピリミジン | G |
二重らせんの 4 つの特徴
| 特徴 | 内容 |
|---|
| ① 2 本鎖 | 2 本のヌクレオチド鎖が互いに巻きつく |
| ② 逆平行 | 1 本が 5'→3'、 もう 1 本が 3'→5' (向きが逆) |
| ③ 相補性 | A と T、 G と C が必ずペア (水素結合) |
| ④ らせん | 10 塩基で 1 周 (約 3.4 nm)、 直径約 2 nm |
大事: シャルガフの規則 (1950): どの生物の DNA でも A の量 = T の量、 G の量 = C の量。 ワトソン・クリックの二重らせんモデルの重要なヒントになりました。
DNA 抽出実験 (バナナ・タマネギ)
家庭でも行える簡単な実験:
- タマネギ や バナナ をすりつぶす
- 食塩水 + 中性洗剤 を加え細胞膜を壊す
- ろ過して上澄みをとる
- 冷やした エタノール をそっと加える
- 境界に 白い糸状の DNA が浮かび上がる
安全配慮: 食塩水・洗剤・エタノールは飲まない、 目に入れない。 エタノール抽出時は 火気厳禁 (引火性)。 抽出した DNA は個人で持ち帰らず、 学校で廃棄します。
4. DNA の複製 — 半保存的複製
細胞分裂の前に、 DNA は 正確に 2 倍 に増えます (複製)。
半保存的複製
1958 年、 メセルソン・スタール が ¹⁵N (重い窒素) で標識した大腸菌の実験で確認:
- 元の二重らせんがほどけ、 各鎖を 鋳型 として新しい鎖が作られる
- できた二重らせんは 1 本が古い鎖、 1 本が新しい鎖 = 半保存的複製
複製に関わる酵素
| 酵素 | 役割 |
|---|
| ヘリカーゼ | 二重らせんをほどく |
| DNAポリメラーゼ | 鋳型に沿って新しいヌクレオチドをつなぐ |
| プライマーゼ | 開始点となる RNA プライマーを作る |
| DNAリガーゼ | 切れた DNA 鎖をつなぐ |
リーディング鎖とラギング鎖
DNAポリメラーゼは 5'→3' 方向にしか 合成できないので、 2 本鎖のうち:
- リーディング鎖: 連続的に合成
- ラギング鎖: 短い断片 (岡崎フラグメント) を何度も作り、 後でDNAリガーゼがつなぐ
大事: 岡崎フラグメントは 1968 年に 岡崎令治・恒子 夫妻 (日本) が発見、 世界的業績。
5. セントラルドグマ — 転写と翻訳
クリックが 1958 年に提唱した 「遺伝情報の流れ」 = セントラルドグマ:
DNA → (転写) → RNA → (翻訳) → タンパク質
転写 — DNA から RNA へ
- 核の中で進行
- DNAの一部がほどけ、 RNAポリメラーゼが鋳型に相補的な mRNA を合成
- DNA の T ↔ RNA では U (ウラシル) に置き換わる
RNA の種類
| 種類 | はたらき |
|---|
| mRNA (伝令 RNA) | 遺伝情報をリボソームへ運ぶ |
| tRNA (転移 RNA) | アミノ酸をリボソームへ運ぶ |
| rRNA (リボソーム RNA) | リボソームを構成 |
翻訳 — RNA からタンパク質へ
- 細胞質の リボソーム で進行
- mRNA を 3 塩基ごと (コドン) に読み、 対応するアミノ酸をtRNAが運んでくる
- アミノ酸がペプチド結合でつながり、 タンパク質になる
遺伝暗号表 — コドン
- 4 種の塩基 × 3 つ = 64 通り のコドンが 20 種のアミノ酸を指定 (重複あり = 縮退)
- 開始コドン AUG = メチオニン
- 終止コドン UAA・UAG・UGA = アミノ酸を指定せず翻訳を終える
| コドン | アミノ酸 |
|---|
| AUG | メチオニン (開始) |
| UUU・UUC | フェニルアラニン |
| UAA・UAG・UGA | (終止) |
真核生物の特徴 — スプライシング
真核生物では、 転写直後の RNA に 不要部分 (イントロン) が含まれます。 これを切り取り、 必要部分 (エキソン) をつなぐ過程が スプライシング。 この後にmRNAとして細胞質へ運ばれます。
6. 突然変異とバイオテクノロジー
突然変異の種類
| 種類 | 内容 | 影響 |
|---|
| 置換 | 1 塩基が別の塩基に変わる | アミノ酸 1 個変化 (同義置換で影響なしも) |
| 挿入 | 塩基が足される | フレームシフトで 以後全てのアミノ酸 がずれる |
| 欠失 | 塩基が抜ける | 同上 |
| 染色体突然変異 | 染色体の一部が入れ替わる・欠ける | ダウン症 (21 番が 3 本) 等 |
大事: 突然変異は 悪いことではない。 多様性を生み、 進化の原動力となる。
遺伝子工学の基礎
| 技術 | 概要 | 応用 |
|---|
| 制限酵素 | DNA を特定の配列で切る | クローニング |
| PCR | DNA を大量に増やす | 親子鑑定・PCR 検査 |
| 電気泳動 | DNA 断片をサイズで分ける | DNA 鑑定 |
| ゲノム編集 (CRISPR-Cas9) | DNA の任意箇所を改変 | 医療・農業 |
7. ふりかえり
この章の安全配慮
- DNA 抽出実験で使う エタノール は火気厳禁、 換気を確保
- 染色液 (酢酸カーミン等) は服に注意、 廃液は容器へ
- 個人の DNA を抽出する実験 (口内細胞) は 同意と個人情報の配慮 を必ず行う
- ゲノム編集等の議論は 生命倫理 を大切にする
次の章: 第 5 章では、 多細胞生物としての 動物の体液調節と恒常性 を学びます。 血液・自律神経・ホルモンがどのように連携して体内を一定に保つかを見ていきましょう。